Livets kjemi drifter proteinmaskiner
Oversatt fra Otangelos FB-side
Det er ingen selektiv fordel før du får den endelige funksjonen.
Lenksy et al. Publiserte en artikkel i Nature 8 som Luskin fra Discovery-instituttet svarte på, hvor igjen Richard B. Hoppe ved 'Pandas thumb' i sin artikkel desperat fremhevet Avida igjen. Debatten er interessant da den berører noen kjernespørsmål om evolusjonen x intelligent design kontroversen.
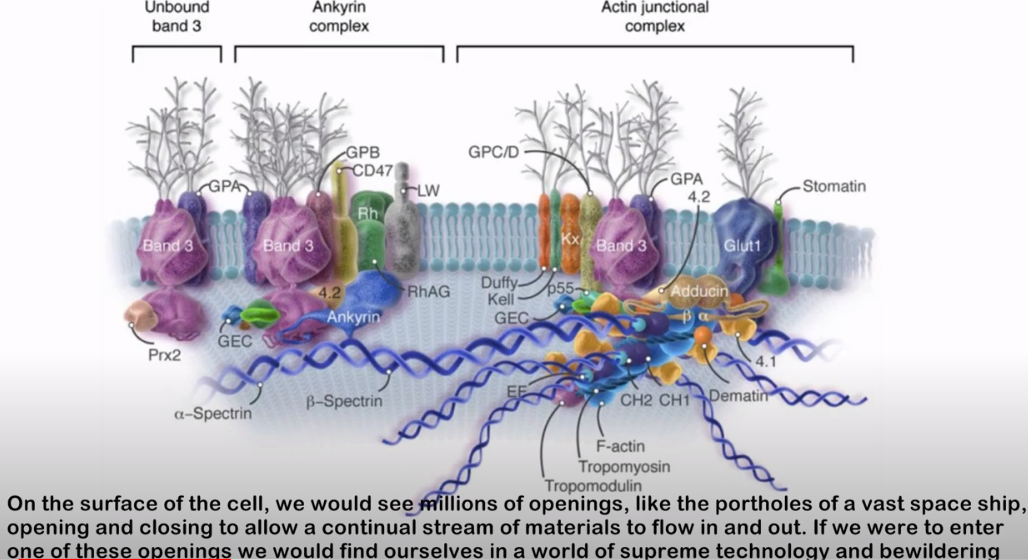 Hoppe skriver følgende som svar til Luskin:
Hoppe skriver følgende som svar til Luskin:
Samvalg og modifisering av eksisterende strukturer er et allestedsnærværende fenomen i evolusjon på nivåer som spenner fra molekylære mekanismer til høynivåstrukturer som vinger.
Dette er en komfortabel måte å unngå de relevante spørsmålene på, og Hoppe unngår dem til tross for at Luskin påpekte at det ikke bare handler om modifisering og samvalg av eksisterende deler, men hvordan de-novo-gener utviklet seg for å starte tilvirkning av nye funksjoner. Hvordan begynte nye strukturer og nye typer celler å utvikle seg?
Biologiske strukturer er godt organisert, strukturert og bygger seg opp som menneskeskapte fabrikker og maskiner. Prosessen foregår helautomatisk inne i cellen. Prosessen med proteinproduksjon, med utgangspunkt i genene, er ekstremt kompleks, og det kreves flere trinn. Bruce Alberts skriver i "The Cell as a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists,":
Men som det viser seg, kan vi gå og vi kan snakke fordi kjemien som gjør livet mulig er mye mer forseggjort og sofistikert enn noe vi noen gang hadde vurdert. Vi vet nå at nesten alle større prosesser i en celle utføres ved samlinger av 10 eller flere proteinmolekyler. Og når den utfører sine biologiske funksjoner, samhandler hver av disse proteinsammensetningene med flere andre store proteinkomplekser. Faktisk kan hele cellen sees på som en fabrikk som inneholder et forseggjort nettverk av samarbeidende samlebånd, hvor hver består av et sett med store proteinmaskiner.
Bestilte bevegelser driver proteinmaskiner
 Hvorfor kaller vi de store proteinsammensetningene som ligger til grunn for cellefunksjonens proteinmaskiner? Nettopp fordi disse proteinsamlingene, i likhet med maskinene oppfunnet av mennesker for å håndtere den makroskopiske verdenen, inneholder sterkt koordinerte bevegelige deler. Innen hver proteinkonstruksjon er intermolekyl ære kollisjoner ikke bare begrenset til et lite sett med muligheter, men reaksjon C avhenger av reaksjon B, som igjen avhenger av reaksjon A - akkurat som den ville gjort i en maskin, etter vår vanlige erfaring.
Hvorfor kaller vi de store proteinsammensetningene som ligger til grunn for cellefunksjonens proteinmaskiner? Nettopp fordi disse proteinsamlingene, i likhet med maskinene oppfunnet av mennesker for å håndtere den makroskopiske verdenen, inneholder sterkt koordinerte bevegelige deler. Innen hver proteinkonstruksjon er intermolekyl ære kollisjoner ikke bare begrenset til et lite sett med muligheter, men reaksjon C avhenger av reaksjon B, som igjen avhenger av reaksjon A - akkurat som den ville gjort i en maskin, etter vår vanlige erfaring.
Under denne høyt organiserte aktiviteten bestilles konformasjonsendringer i ett eller flere proteiner drevet av nukleosid-trifosfathydrolyse (eller av andre energikilder, for eksempel en iongradient). Den nesten allestedsnærværende bruken av energidrevne konformasjonsendringer for å fremme den lokale forsamlingen av proteinkomplekser, og dermed skape en høy grad av orden i cellen, har blitt allment anerkjent.
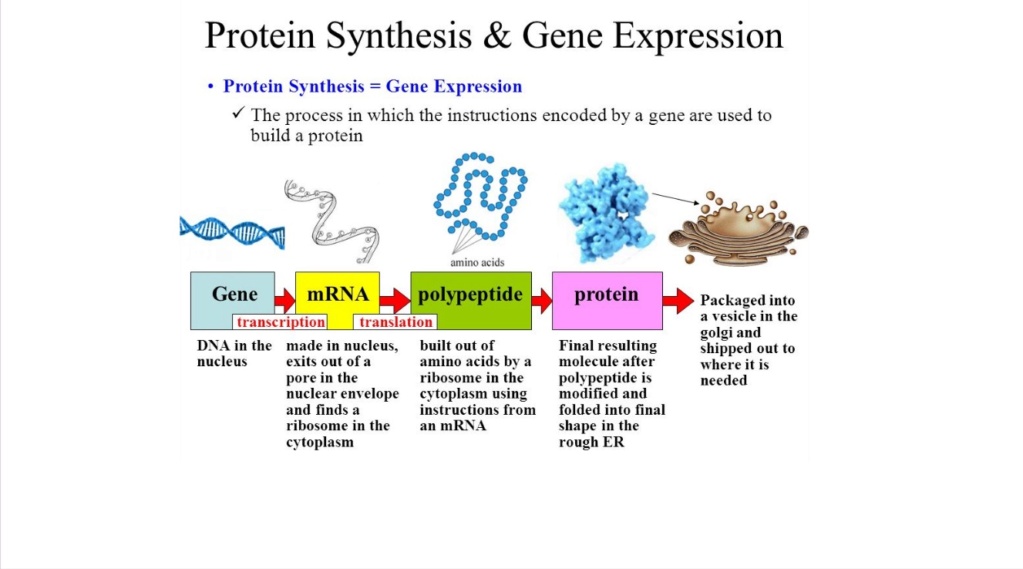
Mange forskjellige typer kjemiske reaksjoner kreves for å produsere et riktig brettet protein fra informasjonen i et gen.
Reisen fra gen til protein er kompleks og tett kontrollert i hver celle. Den består av to hovedtrinn: transkripsjon og oversettelse. Sammen er transkripsjon og oversettelse kjent som genuttrykk.
Cellen sender aktivatorproteiner til stedet for genet som må slås på, som deretter starter RNA-polymerasemaskinen ved å fjerne en plugg som blokkerer DNA-inngangen til maskinen. DNA-strengene skifter posisjon slik at DNA stiller opp med inngangen til RNA-polymerase. Når disse to bevegelsene har skjedd og DNA-strengene er i posisjon, begynner RNA-polymerasemaskinen å smelte dem, slik at informasjonen de inneholder kan behandles for å produsere mRNA 2 Prosessen følger deretter etter INITIASJON AV TRANSKripsjon gjennom RNA-polymeraseenzym komplekser, er mRNA avkortet gjennom post-transkripsjonelle modifikasjoner av flere forskjellige enzymer, ELONGATION gir den viktigste transkripsjonsprosessen fra DNA til mRNA, videre SPLICING og CLEAVAGE, polyadenylering der en lang streng med gjentatte adenosinukleotider blir tilsatt, OG TERMINASJON gjennom over et dusin forskjellige enzymer, EKSPORT FRA KJERNEN TIL CYTOSOLEN (må transporteres aktivt gjennom kjerneporekomplekskanalen i en kontrollert prosess som er selektiv og energiavhengig AV PROTEINSYNTES OG PROTEINFOLDING gjennom chaperone enzymer . Derfra blir proteinene transportert av spesialiserte proteiner til sluttdestinasjonen. De fleste av disse prosessene krever ATP, energidrivstoffet inne i cellen.
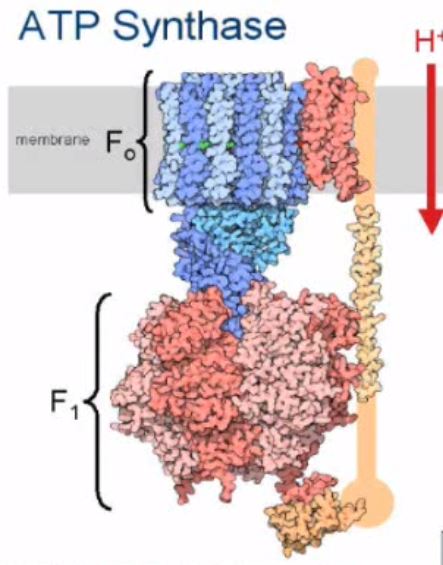 Hvert av disse trinnene krever ekstremt komplekse proteiner og enzymer, arbeidshestene i cellen, som fungerer som roboter i en samlebånd i høyt regulerte presise trinn, og disse maskinene er selv kodet i genomet. Ikke bare er informasjonen for å gjøre dem lagret i genomet. Men disse maskinene krever ytterligere forskjellige proteiner og enzymer for å bli klargjort og satt sammen. Og informasjonen for disse prosessene som foregår er også registrert i genomet. Og noen få gener inneholder informasjonen for å produsere molekyler som hjelper cellen med å samle proteiner, det vil si at oppbyggingen av hele maskineriet også må være forhåndsprogrammert og skje på en sekvensiell, spesiell, ordnet måte. Mange forskjellige prosesser må skje samtidig, drevet av ATP, noe som betyr at ATPase-kraftverket og protongradienten og membranene må være store siden begynnelsen. På samme måte som at vi bygger en maskin, må hver del monteres på rett sted, til rett tid, i riktig rekkefølge og rekkefølge, og delene må passe sammen på en funksjonell og presis måte. Og de riktige materialene er nødvendige. I en bilmotor må stemplene være laget av riktig temperaturbestandige metaller, og slik er det også inne i cellene. De fleste enzymer har reaksjonssentre, der det kreves spesielle substrater og reaktorfaktorer for å utøve sine spesifikke reaksjoner, og mange enzymer krever tilstedeværelse av andre forbindelser - kofaktorer - før deres katalytiske aktivitet kan utøves. 5 Hvordan kunne naturlige teknikker "finne ut" hvilke spesielle materialer, som metall-ion-aktivatorer, som kreves for å gi en gitt reaksjon? Prøving og feiling?
Hvert av disse trinnene krever ekstremt komplekse proteiner og enzymer, arbeidshestene i cellen, som fungerer som roboter i en samlebånd i høyt regulerte presise trinn, og disse maskinene er selv kodet i genomet. Ikke bare er informasjonen for å gjøre dem lagret i genomet. Men disse maskinene krever ytterligere forskjellige proteiner og enzymer for å bli klargjort og satt sammen. Og informasjonen for disse prosessene som foregår er også registrert i genomet. Og noen få gener inneholder informasjonen for å produsere molekyler som hjelper cellen med å samle proteiner, det vil si at oppbyggingen av hele maskineriet også må være forhåndsprogrammert og skje på en sekvensiell, spesiell, ordnet måte. Mange forskjellige prosesser må skje samtidig, drevet av ATP, noe som betyr at ATPase-kraftverket og protongradienten og membranene må være store siden begynnelsen. På samme måte som at vi bygger en maskin, må hver del monteres på rett sted, til rett tid, i riktig rekkefølge og rekkefølge, og delene må passe sammen på en funksjonell og presis måte. Og de riktige materialene er nødvendige. I en bilmotor må stemplene være laget av riktig temperaturbestandige metaller, og slik er det også inne i cellene. De fleste enzymer har reaksjonssentre, der det kreves spesielle substrater og reaktorfaktorer for å utøve sine spesifikke reaksjoner, og mange enzymer krever tilstedeværelse av andre forbindelser - kofaktorer - før deres katalytiske aktivitet kan utøves. 5 Hvordan kunne naturlige teknikker "finne ut" hvilke spesielle materialer, som metall-ion-aktivatorer, som kreves for å gi en gitt reaksjon? Prøving og feiling?
Videre kreves følgende:
C1: Tilgjengelighet. Blant delene som er tilgjengelige for rekruttering for å danne et biologisk system bestående av flere deler, må det være noen som kan utføre de høyt spesialiserte oppgavene til det spesifikke systemet, selv om alle elementene tjener en annen funksjon eller ingen funksjon i et annet system hvor de ble rekruttert fra.
C2: Synkronisering. Tilgjengeligheten av disse delene må synkroniseres slik at de på et eller annet tidspunkt, hver for seg eller i kombinasjon, er tilgjengelige samtidig.
C3: Lokalisering. De valgte delene må alle gjøres tilgjengelige på samme 'byggeplass', kanskje ikke samtidig, men absolutt på det tidspunktet de er nødvendige.
C4: Koordinering. Delene må være gjensidige kompatible, det vil si "godt matchet" og i stand til å 'samhandle' riktig: selv om underenhetene er satt sammen i riktig rekkefølge, må de også ha rett grensesnitt . Delene må koordineres på riktig måte: selv om alle delene av et ribosom er tilgjengelig til rett tid, er det klart at de fleste måter å montere dem på vil være ikke-funksjonelle eller irrelevante.
C5: Grensesnittkompatibilitet. Delene må være gjensidige kompatible, det vil si 'godt matchet' og i stand til å 'samhandle ordentlig': selv om underenhetene er satt sammen i riktig rekkefølge, må de også fungere riktig i grensesnittet.
Så disse ytterligere spørsmålene oppstår:
Av hvilken grunn ville naturlige prosesser produsere maskinene som for eksempel Ribonuclease P, som behandler pre-tRNA, som inneholder ekstra tRNA-sekvenser i både 5’- og 3’-endene og må fjernes? Av hvilken grunn ville naturlige prosesser produsere tRNA som kreves inne i Ribosomet, det sentrale molekylet i oversettelsesprosessen?
P Ribonuklease P ville ikke ha noen funksjon alene. tRNA har ingen funksjon alene. Ribosomet har ingen funksjon alene. Disse individuelle delene utøver bare sin funksjon, hvis de er griper inn i hverandre, og arbeider på en gjensidig avhengig måte sammen. Anta at alt ville starte gjennom naturlige prosesser, hvordan kunne disse maskinene oppstå hver for seg, på en trinnvis måte, hvis de ikke har noe eget bruk? Det er ikke bare at 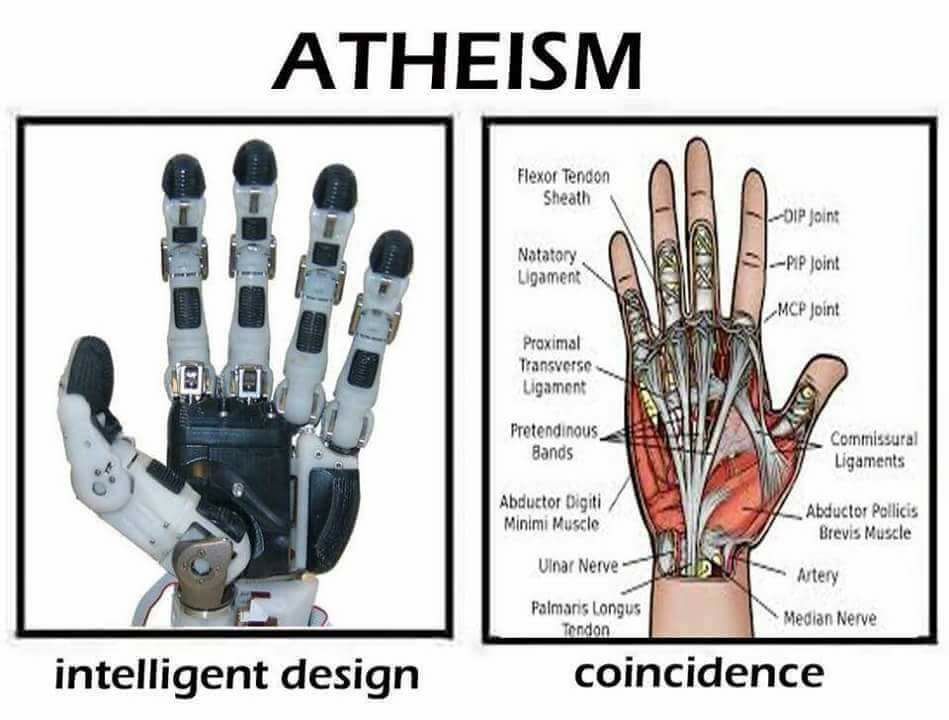 vi kan argumentere for at vi ikke vet det ennå. Det vi vet, tillater oss rasjonelt å utlede at naturalistiske forklaringer er helt utilstrekkelige til å forklare de aktuelle fenomenene. En innledende tegning er nødvendig, der hele prosessen er forhåndsprogrammert, noe som er tilfelle i genomet, der all informasjonen for å bygge en celle er lagret, og hele prosessen må starte på en gang. Det ser ut til å forklares best gjennom en intelligent designer.
vi kan argumentere for at vi ikke vet det ennå. Det vi vet, tillater oss rasjonelt å utlede at naturalistiske forklaringer er helt utilstrekkelige til å forklare de aktuelle fenomenene. En innledende tegning er nødvendig, der hele prosessen er forhåndsprogrammert, noe som er tilfelle i genomet, der all informasjonen for å bygge en celle er lagret, og hele prosessen må starte på en gang. Det ser ut til å forklares best gjennom en intelligent designer.
Slik 'Uncommondescent' uttrykker det:
ID foreslår ikke at 'Gud' skal skrive over et gap i den nåværende vitenskapelige forklaringen. I stedet begynner ID-teoretikere fra empirisk observerte, pålitelige, kjente fakta og allment aksepterte prinsipper for vitenskapelig resonnement:
(a) Intelligente designere eksisterer og handler i verden.
(b) Når de gjør det, etterlater de som regel pålitelige tegn på en slik intelligent handling.
(c) Faktisk, for mange av de aktuelle tegnene som CSI og IC, er intelligente agenter den eneste observerte årsaken til slike effekter, og tilfeldighet + nødvendighet (alternativet) er ikke en sannsynlig kilde, fordi funksjonelle 'øyer' er mye for sjeldne i mulige relevante konfigurasjoner.
(d) På grunnlag av vitenskapens generelle prinsipp, at 'lik årsaker som', har vi derfor rett til å utlede fra tegn til betydning: intelligent handling.
(e) Denne konklusjonen er selvfølgelig gjenstand for falsifisering, hvis det kan vises at ikke-styrt tilfeldighet + mekaniske krefter gir CSI eller IC. Dermed er ID falsifiserbart i prinsippet, men godt støttet i realiteten.
I sum er ID faktisk en legitim vitenskapelig innsats: vitenskapen som studerer tegn på intelligens.
Oversettelse og bilder ved Asbjørn E. Lund